Varalu is an early maturing rice variety widely grown in the rainfed ecosystem preferred for its grain type and cooking quality. However, the yield of Varalu is substantially low since it is being affected by reproductive drought stress along with the blast disease. The genetic improvement of Varalu was done by introgressing a major yield QTL, qDTY12.1, along with two major blast resistance genes i.e. Pi54 and Pi1 through marker-assisted backcross breeding. Both traits were transferred till BC2 generation and intercrossing was followed to pyramid the two traits. Stringent foreground selection was carried out using linked markers as well as peak markers (RM28099, RM28130, RM511 and RM28163) for the targeted QTL ( qDTY12.1), RM206 for Pi54 and RM224 for Pi1. Extensive background selection was done using genome-wide SSR markers. Six best lines (MSM-36, MSM-49, MSM-53, MSM-57, MSM-60 and MSM-63) having qDTY12.1 and two blast resistance genes in homozygous condition with recurrent parent genome of 95.0%‒96.5% having minimal linkage drag of about 0.1 to 0.7 Mb were identified. These lines showed yield advantage under drought stress as well as irrigated conditions. MSM-36 showed better performance in the national coordinated trials conducted across India, which indicated that improved lines of Varalu expected to replace Varalu and may have an important role in sustaining rice production. The present study demonstrated the successful marker-assisted pyramiding strategy for introgression of genes/QTLs conferring biotic stress resistance and yield under abiotic stress in rice.
Rice (Oryza sativa L.) is a primary source of nutrition for more than half of the world’ s population. Despite the saturation in genetic yield, demand for rice is increasing with the ever increasing population. Rice is also cultivated in around 23 million hectares of rainfed ecosystem where yields are significantly affected due to frequent occurrence of drought (Serraj et al, 2011), and blast (Dean et al, 2012), and global climate change can further increase the drought stress (Wassmann et al, 2009). Also, yield is affected due to drought spells in irrigated conditions (Bernier et al, 2009a).
Among many QTLs identified for drought stress tolerance, qDTY12.1offers significant yield potential under reproductive-stage drought stress by contributing 51% genetic variance (Bernier et al, 2007). Blast disease caused by Magnaporthe grisea(anamorph Pyricularia grisea) is another production constraint commonly observed in the rainfed ecosystem (Dean et al, 2012). So far, above 100 genes have been reported to aid in blast resistance (Devi et al, 2020). Among them, two significant blast resistance genes Pi54and Pi1 confer broad-spectrum resistance against predominant races of the pathogen in India (Mackill and Bonman, 1992; Sharma et al, 2005). Marker-assisted gene pyramiding (MAGP) is a precise, rapid and efficient method to pyramid desired genes/QTLs along with maximum recovery of recurrent parent genome within two to three backcrosses (Jamaloddin et al, 2020).
Varalu is a popular variety cultivated in major upland areas of India. It is affected by reproductive drought stress and blast disease. Hence, this study aimed to genetic improvement of Varalu by introgressing a major yield QTL (qDTY12.1) along with two major blast resistance genes (Pi54 andPi1) through MAGP.
RESULTS
Introgression ofqDTY12.1 into Varalu
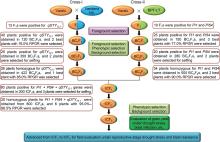
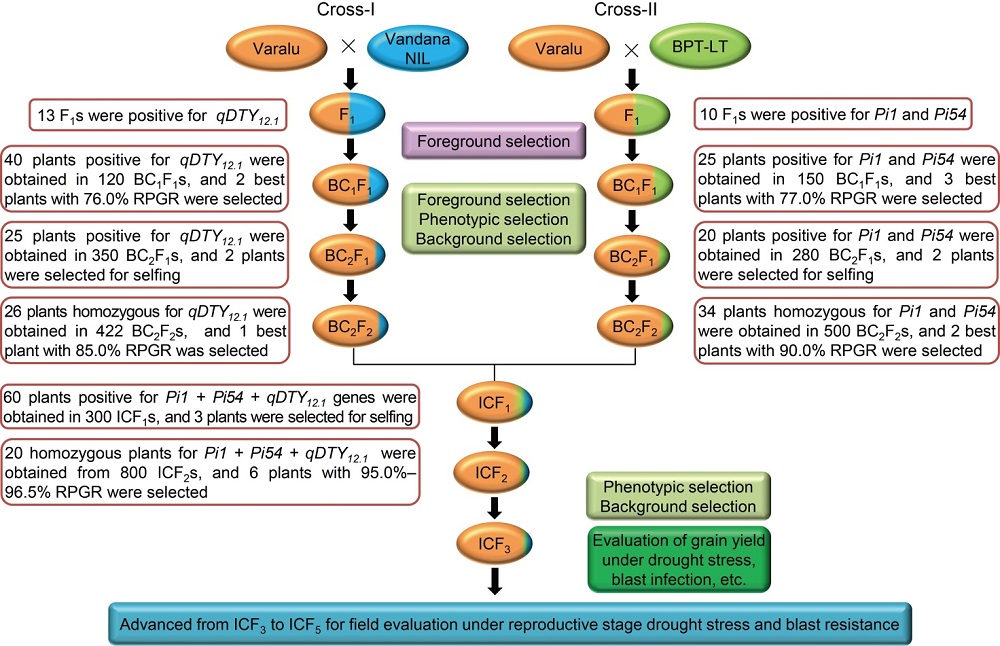
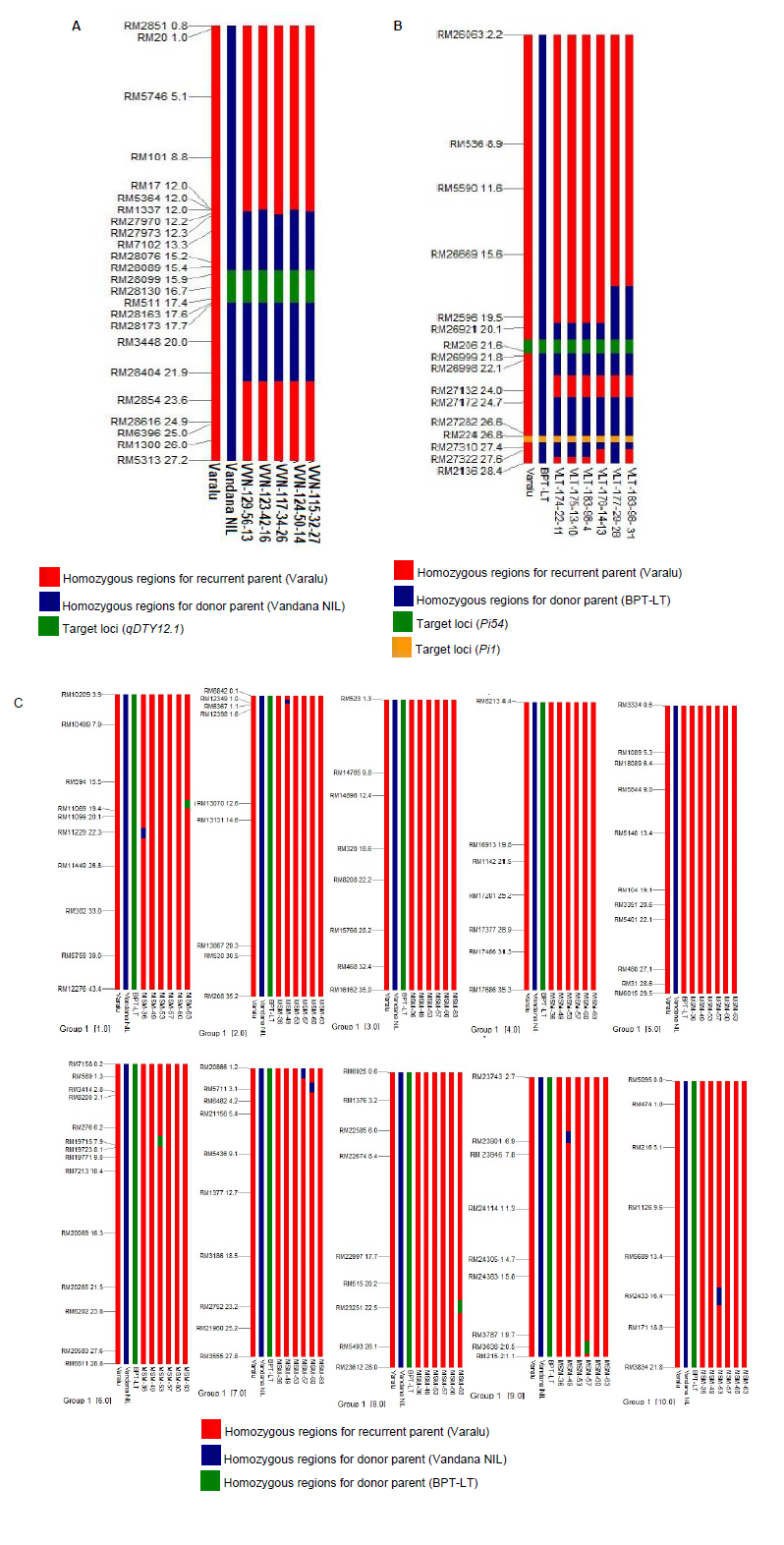
Through foreground selection, 13 F1 plants from Cross-I [Varalu × Vandana near-isogenic line (NIL)], 40 BC1F1, 25 BC2F1and 26 BC2F2plants were found positive for qDTY12.1 (Fig. 1). Polymorphic assay between Varalu and Vandana NIL identified 200 SSR markers, which were uniformly spread across the rice genome. Two best BC1F1plants possessing the maximum recurrent parent genome recovery (RPGR, 76.0%) and qDTY12.1 were backcrossed to produce BC2F1, and two best BC2F1 plants were used to produce BC2F2 population (n = 422). Finally, five BC2F2 plants displaying homozygous qDTY12.1 genomic region and high grain yield under reproductive drought stress portraying 82.0%‒85.0% of RPGR were identified. The recombinant selection was performed among five BC2F2 plants to minimize the linkage drag at qDTY12.1 locus and found the recombination breakpoints (RBP) ranged from 3.6 to 4.3 Mb at the proximal and distal ends, respectively (Fig. S1-A). Among them, one plant (VVN-117-34-26) with the maximum RPGR of nearly 85.0% was used for intercrossing.
Fig. 1. Schematic representation of marker-assisted gene pyramiding followed for introgression of qDTY12.1 for improvement of grain yield under reproductive stage drought stress and blast resistance genes (Pi54 and Pi1) in Varalu variety. NIL, Near-isogenic line; BPT-LT, Elite rice line with the genetic background of Samba Mahsuri containing two blast resistance genes Pi54and Pi1; RPGR, Recurrent parent genome recovery; IC, Intercross.
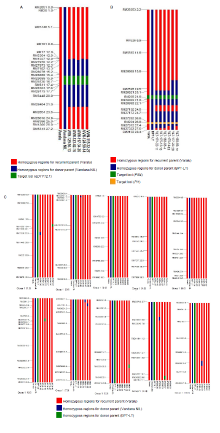
Fig. S1. Graphical representation of BC2F2 selected lines. A, Graphical representation of BC2F2 selected lines of cross-I (Varalu × Vandana NIL) for the donor genome introgression associated with the qDTY12.1 on target chromosome 12. Red colour indicates homozygous regions for recurrent parent Varalu, blue colour indicates the donor parents (BPT-LT) and green colour indicates qDTY12.1. =B, Graphical representation of BC2F2 selected lines of cross-II (Varalu × BPT-LT) for the donor genome introgression associated with the blast resistance genes, Pi54 and Pi1 on target chromosome 11. Red colour indicates homozygous regions for recurrent parent Varalu, blue colour indicates the donor parents (BPT-LT), green colour indicates Pi54 gene and black colour indicatesPi1 gene. C, Graphical representation of pyramided lines for the donor genome introgression on non-target chromosomes 1‒10. Red colour indicates homozygous regions for recurrent parent Varalu and the green and blue colours indicate the donor parents (BPT-LT and Vandana NIL) genome region, respectively.
Introgression of blast resistance genes into Varalu
Ten F1 plants derived from Cross-II [Varalu × BPT-LT (an elite rice line with the genetic background of Samba Mahsuri containing two blast resistance genes Pi54and Pi1)], 25 BC1F1, 20 BC2F1 and 34 BC2F2 plants were found positive for both Pi54 and Pi1 (Fig. 1). A total of 180 markers showed polymorphism between Varalu and BPT-LT were used for background analysis. Three best BC1F1 plants possessing a maximum RPGR of 77.0% were used to generate BC2F1 plants and two best BC2F1 plants were used to produce BC2F2 plants. Finally, six BC2F2plants showed a high resistant level to blast with 86.0%‒90.0% of RPGR. The RBP of six plants at Pi54 locus was observed at 2.1 to 0.5 Mb of donor genome segment at proximal and distal ends, respectively (Fig. S1-B). In case of Pi1 locus, all the six plants showed RBP of 2.1 Mb at the proximal end, and ranged from 0.6 to 1.6 Mb at the distal end. Two BC2F2 plants (VLT-175-13-10 and VLT-183-98-31) having Pi54 and Pi1 in homozygous condition with the maximum RPGR of nearly 90.0% were forwarded for intercrossing (IC).
Pyramiding of qDTY12.1 and blast resistance genes into Varalu
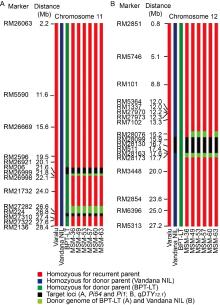
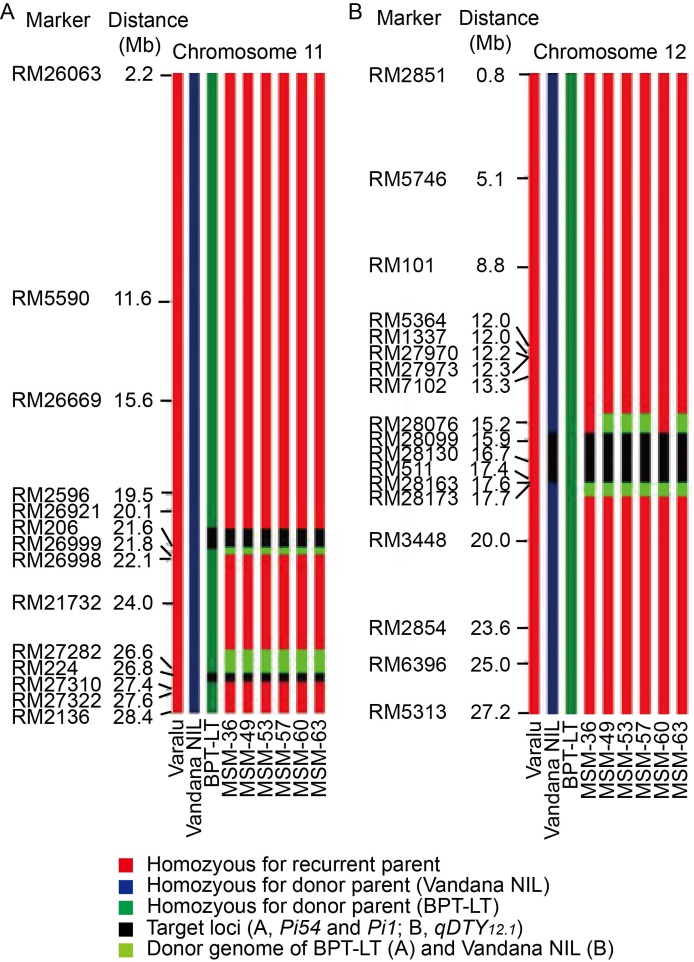
The best homozygous BC2F2 plants obtained from the two crosses were intercrossed for pyramiding the target genes/QTL (qDTY12.1, Pi54and Pi1). The best three ICF1 plants having all the three genes were selfed to generate ICF2 population. Among these, 20 ICF2 plants were homozygous for all the three genes (qDTY12.1, Pi54and Pi1). Six best pyramided lines (MSM-36, MSM-49, MSM-53, MSM-57, MSM-60 and MSM-63) were identified based on blast resistance and grain yield under reproductive stage drought stress (Fig. 1). Background analysis with 120 polymorphic SSR markers revealed 95.0%‒96.5% of RPGR (Fig. S1-C). The recombinant selection among the six best lines revealed a segment of 0.3 and 0.2 Mb of donor genome segment at the distal and proximal ends of Pi54 and Pi1 loci, respectively (Fig. 2-A). Moreover, atqDTY12.1 locus, 0.7 and 0.1 Mb of donor genome segment was found at the proximal and distal ends, respectively (Fig. 2-B). Among the best six lines, MSM-36 and MSM-60 showed very less linkage drag in the three loci compared to the other four entries.
Fig. 2. Graphical representation of selected pyramided lines of Varalu for donor genome introgression associated with blast resistance genes Pi54 and Pi1 on chromosome 11 and qDTY12.1 on chromosome 12. A, At Pi54 locus, a donor segment introgression was limited to only about 0.3 Mb at the proximal end; at Pi1 locus, about 0.2 Mb donor genome was observed at the proximal end. B, At qDTY12.1 locus, a donor genomic region about 0.7 and 0.1 Mb at the proximal and distal ends, respectively, was observed in four lines on chromosome 12. Two lines (MSM-36 and MSM-60) showed limited donor segments in comparison with other lines. NIL, Near-isogenic line; BPT-LT, Elite rice line with the genetic background of Samba Mahsuri containing two blast resistance genes Pi54and Pi1.
Performance of improved lines for yield under drought and irrigated conditions
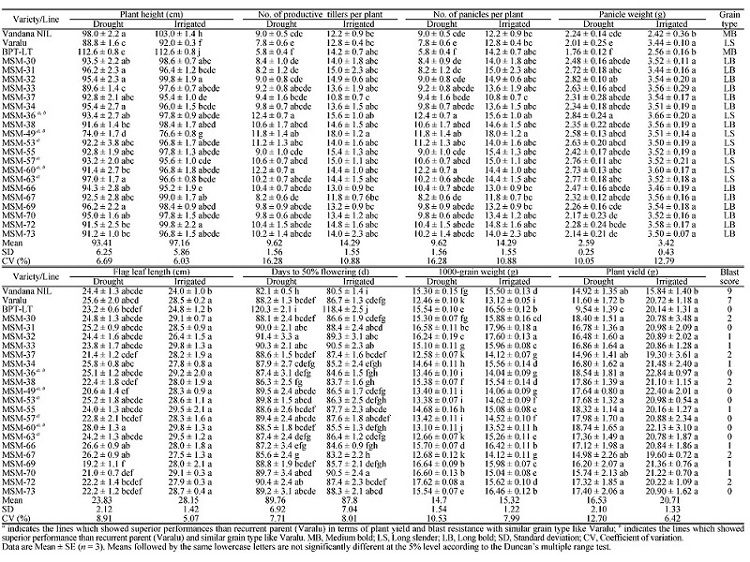
Among the 422 BC2F2 individuals of Cross-I, 26 homozygous plants containing qDTY12.1 with higher yields ranging from 14.8 to 23.4 g/plant, which was chosen for intercrossing. Twenty of the 800 ICF2s displayed higher grain yield per plant (14.96 to 18.74 g) than the recurrent parent (RP) (11.6 g) under reproductive stage drought stress (Table 1). Most of the intercrossed lines had higher plant height except MSM-49, which was shorter than Varalu (Table 1 and Fig. S2-A). Interestingly, the measured yield contributing traits were also superior to RP in these lines (Table 1 and Fig. S2-B). However, there was little difference in the flag leaf length and days to 50% flowering among the improved lines (Table 1). Among these, six improved lines (MSM-36, MSM-49, MSM-53, MSM-57, MSM-60 and MSM-63) having superior yield characteristics and similar grain type like RP in reproductive stage drought stress conditions were selected and further assessed for yield under irrigated conditions (Table 1; Fig. S2-B and -C).
Table 1
Table 1
Table 1 Agronomic performance of pyramided lines of Varalu under reproductive stage drought stress and irrrigated conditions.
Table 1 Agronomic performance of pyramided lines of Varalu under reproductive stage drought stress and irrrigated conditions.
Fig. S2. Phenotypes of improved lines. A, Representation of single plants of improved lines (Pi54 + Pi1 + qDTY12.1) and six selected improved lines of Varalu, showing superior performance in comparison with the recurrent parent under drought conditions. B, Panicles of the recurrent parent and selected improved lines of Varalu. C, Grain type of selected lines showing similarity with the recurrent parent.
Under irrigated conditions, three lines, MSM-36, MSM-49 and MSM-60, exhibited higher grain yield (22.84, 22.40 and 22.13 g, respectively) than the recurrent parent (20.72 g) (Table 1).
Evaluation of disease reaction in improved lines of Varalu
The six BC2F2 plants of Cross-II and 20 homozygous pyramided lines ICF2to ICF5 noted ‘ 0’ (Fig. S3) and ‘ 0‒2’ blast scores, respectively (Table 1 and Fig. 3), indicating high level of blast resistance.
Fig. 3. Phenotypic screening of pyramided lines (Pi54 + Pi1 + qDTY12.1) against blast disease. A, All the intercross derived lines and donor parent BPT-LT (an elite rice line with the genetic background of Samba Mahsuri containing two blast resistance genes Pi54and Pi1) were highly resistant whereas the recurrent parent Varalu showed susceptible against blast. B, Lesions were observed on the leaf surface of the recurrent parent Varalu while the pyramided lines and donor parent showed no lesion on the leaf surface.
Moreover, MSM-36 out yielded its RP in Zone VII of All India Coordinated Rice Improvement Programme (AICRIP, consisting of five states, Telangana, Andhra Pradesh, Karnataka, Tamil Nadu and Kerala) with yield advantage of 18.8% and 13.8% under drought and controlled conditions, respectively. The average blast disease susceptibility index of MSM-36 at multiple locations was 2.95 compared to Varalu of 6.20 (Table S2).
Table S2
Table S2
Table S2 Performance of best pyramided lines under drought and controlled conditions in national trials (Zone VII).
Table S2 Performance of best pyramided lines under drought and controlled conditions in national trials (Zone VII).
Grain quality analysis of improved lines
In addition to the brown hull colour, long slender grain type and cooking quality parameters of the six pyramided lines were assessed and found MSM-36 and MSM-60 were particularly similar to RP (Table 2).
Table 2
Table 2
Table 2 Grain and cooking quality of selected pyramided lines of Varalu.
Variety/Line
Hulling rate (%)
Milling rate (%)
HRR (%)
GL (mm)
GW (mm)
GL/GW
Grain type
Grain chalkiness
ASV
AC (%)
GC (mm)
Varalu
77.9
68.9
57.3
6.13
2.04
3.04
LS
VOC
7
25.51
28
Vandana NIL
66.7
59.3
52.4
5.90
2.20
2.68
MB
VOC
3
23.12
63
BPT-LT
79.6
71.2
68.5
5.40
2.10
2.57
MB
VOC
4
27.61
61
MSM-36
78.9
69.5
59.8
6.12
2.05
3.00
LS
VOC
7
26.79
27
MSM-49
77.9
69.8
58.2
6.04
2.01
3.00
LS
VOC
4
25.34
28
MSM-53
77.5
68.5
58.3
6.10
2.02
3.02
LS
VOC
6
26.38
26
MSM-57
78.7
68.4
59.2
6.12
2.02
3.04
LS
VOC
7
26.18
28
MSM-60
78.2
69.4
58.7
6.17
2.03
3.04
LS
VOC
7
25.88
29
MSM-63
78.4
68.6
59.7
6.11
2.03
3.01
LS
VOC
6
26.58
29
HRR, Head rice recovery; GL, Grain length; GW, Grain width; GL/GW, Ratio of grain length to width; VOC, Very occasionally; ASV, Alkali spreading value; AC, Amylose content; GC, Gel consistency; LS, Long slender; MB, Medium bold.
Table 2 Grain and cooking quality of selected pyramided lines of Varalu.
DISCUSSION
Several drought-tolerant QTLs detected from diverse sources (Kumar et al, 2007; Vikram et al, 2011) are less exploited in breeding programmes. There is a keen interest in selecting the progenies having higher yield even under the stress, which is offered by a few QTLs like qDTY12.1 (Bernier et al, 2009b). The effectiveness of qDTY12.1 was evident from the introgressions of Vandana and Sabitri under drought and irrigated conditions (Mishra et al, 2013; Kumar et al, 2014). Severe incidence of blast disease was also seen in the upland ecosystem (Asibi et al, 2019), which can be addressed by pyramiding major blast resistance genes.
Marker-assisted breeding strategy has been employed to improve rice varieties for resistance against biotic and biotic stresses independently (Dixit et al, 2020). But few reports exist on the development of combined resistant versions to biotic and abiotic stress resistant varieties (Dixit et al, 2014, 2020; Muthu et al, 2020).
RPGRs of the two backcrosses (Cross-I and Cross-II) and intercross were 82.0%‒85.0%, 86.0%‒90.0% and 95.0%‒96.5%, respectively, which were higher than the reported introgression studies (Khan et al, 2018; Chukwu et al, 2019; Swathi et al, 2019). Moreover, the improved lines displayed very close recombination breakpoints at both ends of the three loci, indicating less donor genome introgression. Anyaoha et al (2019) improved the rainfed rice variety FUNAABOR-2 for grain yield under drought stress by introgressing qDTY12.1 and qDTY2.3 using marker-assisted selection with the minimum linkage drag of about 0.8 Mb. Like Pi54 and Pi1pyramided Tellahamsa lines (Jamaloddin et al, 2020), individuals of backcross population having both the genes noted higher resistance score of 0 than the individuals with only Pi54 (0‒2) or Pi1 (2‒3). Thus, Pi54 might provide broad spectrum resistance than Pi1(Patroti et al, 2019). The NILs possessing two genes or single gene can be used as donors for the deployment in the rice improvement programme. Like Sabitri lines under lowland reproductive stage drought stress (Mishra et al, 2013), the backcross population having properly delimited qDTY12.1 A+B region of Dixit et al (2012) showed higher grain yield than the lines possessing either one.
Due to proper phenotypic selection, the mean grain yields of pyramided lines under drought (16.53 g) and irrigated (20.71 g) conditions were higher than the RP (11.60 g and 20.72 g) (Table 1).The results indicated that agro-morphological and cooking quality characters of the improved lines of Varalu were similar to RP except MSM-49 with shorter plant height and lower alkali spreading value. Under the reproductive stage drought stress and irrigated conditions, MSM-36 and MSM-60 were identified as the best lines with high grain yield and desirable cooking quality (Table 2), and MSM-36 noted superior performance in the Zone VII of AICRIP trial.
The present study combined a major drought QTL (qDTY12.1) and two major blast resistance genes (Pi54 and Pi1) into the genetic background of upland rice variety Varalu. Two best improved Varalu lines, MSM-36 and MSM-60 possessing high yield under drought stress and blast resistance along with > 96% RPGR are expected to replace Varalu. These lines did not show any yield penalty under irrigated condition and can be disseminated for cultivation in drought- prone niches of India.
METHODS
Rice materials
The long slender and early maturing (90‒95 d) elite rice variety, Varalu (WGL-20471 × CR-544-1-2), was used as the recurrent parent. Vandana NIL and BPT-LT were used as donors for drought (qDTY12.1) (Bernier et al, 2007) and blast (two resistance genes Pi54 and Pi1), respectively. Two independent crosses viz. Varalu × Vandana NIL (Cross-I) and Varalu × BPT-LT (Cross-II) were made followed by selection of positive lines and backcrossing with Varalu up to BC2F2 and intercrossing at BC2F2 for pyramiding both traits. The selected lines possessing qDTY12.1 + Pi54 + Pi1were advanced through the pedigree method from ICF3 to ICF5 for field evaluation of agronomical traits against blast and reproductive stage drought stress (Fig. 1).
DNA extraction and genotyping
Genomic DNA was isolated from 21-day-old seedlings of the parents, F1, backcross and intercross generations (Murray and Thompson, 1980). Devi et al (2015)procedure was used for PCR amplification. The foreground selection of qDTY12.1 was done using peak marker as well as linked markers RM511, RM28099 (for qDTY12.1 A), RM28130 and RM28163 (for qDTY12.1 B), while RM206 for Pi54, RM224 for Pi1 (Dixit et al, 2012; Patroti et al, 2019) (Table S1.). A set of 635 SSR markers covering every 2‒5 Mb interval were selected (http://rice. plantbiology.msu.edu/) for background selection. The identified polymorphic markers between the donors and the recurrent parent were utilized for background selection in backcross and intercross populations. The assessment of RPGR was done based on the SSR marker data using the Graphical Genotypes (GGTs) V2.0 software.
Table S1
Table S1
Table S1 Details of SSR markers used for foreground selection.
Chr. Chromosome; DP, Donor parenr; RP, Recurrent parent.
Table S1 Details of SSR markers used for foreground selection.
Screening of derived lines under reproductive stage drought stress
Backcross and intercross derived populations were screened for reproductive stage drought stress (Venuprasad et al, 2007) at the research farm, ICAR-Indian Institute of Rice Research (IIRR), Hyderabad, India, in a random block design with 20 cm × 20 cm row spacing. Agro-morphological traits like plant height, number of tillers per plant, number of panicles per plant, flag leaf length, days to 50% flowering, 1000-grain weight and grain yield were measured.
Screening for blast
The blast screening of all the backcross and intercross lines of Varalu was done in a uniform blast nursery at ICAR-IIRR (Umakanth et al, 2017) using Standard Evaluation System with 0‒9 scales (IRRI, 2002).
Screening of improved lines under irrigated conditions
Twenty-five-day-old seedlings of selected lines along with the parents were transplanted in the lowland irrigated plot in a random block design with 150 cm × 20 cm row spacing. Irrigation was given twice a week and all the agro- morphological traits measured in drought plot were also recorded. Further, two best lines (MSM-36 and MSM-60) were also screened across 10 different AICRIP locations under reproductive stage drought stress and irrigated conditions (http://www.icar-iirr.org/aicrip.htm).
Quality analysis
The harvested seeds of the selected lines in triplicate and their parents were shade dried up to 12%‒14% moisture content. Intact milled grains were used to determine grain type by length to width ratio (IRRI, 2004) and alkali spreading value (Juliano et al, 1990). Milled grains ground to 100 mesh powders were analyzed for gel consistency (Cagampang, 1973) and amylose content (Juliano, 1971).
Statistical analysis
The Duncan’ s multiple range test was carried out to compare significant difference of improved lines with recurrent parent for agro-morphological traits using XLSTAT (Version 2020.5). The coefficient of variation, standard deviation and standard error of the mean were calculated by using the MS Excel package to determine the significant variation between the lines.
ACKNOWLEDGEMENT
The authors acknowledge the Department of Biotechnology, New Delhi, India for providing funds for carrying out the research work.
AnyaohaC O, FofanaM, GracenV, TongoonaP, Mand eS. 2019. Introgression of two drought QTLs into FUNAABOR-2 early generation backcross progenies under drought stress at reproductive stage. Rice Sci, 26(1): 32‒41. [Cited Within:1]
[2]
AsibiA E, ChaiQ, CoulterJ A. 2019. Rice blast: A disease with implications for global food security. Agronomy, 9(8): 451. [Cited Within:1]
[3]
BernierJ, KumarA, VenuprasadR, SpanerD, AtlinG N. 2007. A large-effect QTL for grain yield under reproductive-stage drought stress in upland rice. Crop Sci, 47(2): 507‒516. [Cited Within:2]
[4]
BernierJ, SerrajR, KumarA, VenuprasadR, ImpaS, GowdaV R, OaneR, SpanerD, AtlinG. 2009a. The large-effect drought- resistance QTL qtl12. 1 increases water uptake in upland rice. Field Crops Res, 110: 139‒146. [Cited Within:1]
[5]
BernierJ, KumarA, VenuprasadR, SpanerD, VerlukarS, Mand alN P, SinhaP K, PeerajuP, DongreP R, MahtoR N, AtlinG. 2009b. Characterization of the effect of a QTL for drought resistance in rice, qtl12. 1 over a range of environments in the Philippines and eastern India. Euphytica, 166(2): 207‒217. [Cited Within:1]
[6]
CagampangG B, PerezC M, JulianoB O. 1973. A gel consistency test for eating quality of rice. J Sci Food Agric, 24(12): 1589‒1594. [Cited Within:1]
[7]
ChukwuS C, RafiiY M, RamleeS I, IsmailS I, OladosuY, OkporieE, OnyishiG, UtoboE, EkwuL, SwarayS, JallohM. 2019. Marker-assisted selection and gene pyramiding for resistance to bacterial leaf blight disease of rice ( Oryza sativa L. ). Biotechnol Biotechnol Equip, 33(1): 440-455. [Cited Within:1]
[8]
DeanR, van Kan J A L, PretoriusZ A, Hammond-KosackK E, DiPietro A, SpanuP D, RuddJ J, DickmanM, KahmannR, EllisJ, FosterG D. 2012. The Top 10 fungal pathogens in molecular plant pathology. Mol Plant Pathol, 13(4): 414-430. [Cited Within:2]
[9]
Devi S J S R, KuldeepS, UmakanthB, VishalakshiB, RenukaP, SudhakarK V, PrasadM S, ViraktamathB C, RavindrababuV, MadhavM S. 2015. Development and identification of novel rice blast resistant sources and their characterization using molecular markers. Rice Sci, 22(6): 300-308. [Cited Within:1]
[10]
Devi S J S R, SinghK, UmakanthB, VishalakshiB, RaoK V S, SuneelB, SharmaS K, KadambariG K M, PrasadM S, SenguttvelP, DivyaP S, MadhavM S. 2020. Identification and characterization of a large effect QTL from Oryza glumaepatula revealed Pi68(t) as putative cand idate gene for rice blast resistance. Rice, 13: 17. [Cited Within:1]
[11]
DixitS, SwamyB M, VikramP, AhmedH U, CruzM S, AmanteM, AtriD, LeungH, KumarA. 2012. Fine mapping of QTLs for rice grain yield under drought reveals sub-QTLs conferring a response to variable drought severities. Theor Appl Genet, 125(1): 155-169. [Cited Within:2]
[12]
DixitS, HuangB E, StaCruz M T, MaturanP T, OntoyJ C E, KumarA. 2014. QTLs for tolerance of drought and breeding for tolerance of abiotic and biotic stress: An integrated approach. PLoS One, 9(10): e109574. [Cited Within:1]
[13]
DixitS, SinghU M, SinghA K, AlamS, VenkateshwarluC, NachimuthuV V, YadavS, AbbaiR, SelvarajR, DeviM N, RamayyaP J, BadriJ, RamT, LakshmiJ, LakshmideviG, Vidhya L R K J, PadmakumariA P, LahaG S, PrasadM S, SeetalamM, SinghV K, KumarA. 2020. Marker assisted forward breeding to combine multiple biotic-abiotic stress resistance/tolerance in rice. Rice, 13: 29. [Cited Within:2]
[14]
IRRI. 2002. Stand ard Evaluation System for Rice. Los Banos, Manila, the Philippine: International Rice Research Institute. [Cited Within:1]
[15]
IRRI. 2004. Stand ard Evaluation System for Rice. Los Banos, Manila, the Philippine: International Rice Research Institute. [Cited Within:1]
[16]
JamaloddinM, DurgaRani C V, SwathiG, AnuradhaC, VanisriS, RajanC P D, KrishnamRaju S, BhuvaneshwariV, JagadeeswarR, LahaG S, PrasadM S, SatyanarayanaP V, CheraluC, RajaniG, RamprasadE, SravanthiP, ArunPrem Kumar N, ArunaKumari K, YaminiK N, MaheshD, SanjeevRao D, SundaramR M, SheshuMadhav M. 2020. Marker assisted gene pyramiding (MAGP) for bacterial blight and blast resistance into mega rice variety ‘Tellahamsa’. PLoS One, 15(6): e0234088. [Cited Within:2]
[17]
JulianoB O. 1971. A simplified assay for milled rice amylose. Cereal Sci Today, 16(11): 334-360. [Cited Within:1]
[18]
JulianoB O, PerezC M, KaushikR, KhushG S. 1990. Some grain properties of IR36-based starch mutants. Starch, 42(7): 256-260. [Cited Within:1]
[19]
KhanG H, ShikariA B, VaishnaviR, NajeebS, PadderB A, BhatZ A, ParrayG A, BhatA M, KumarR, SinghN K. 2018. Marker-assisted introgression of three dominant blast resistance genes into an aromatic rice cultivar Mushk Budji. Sci Rep, 8(1): 4091. [Cited Within:1]
[20]
KumarA, DixitS, RamT, YadawR B, MishraK K, Mand alN P. 2014. Breeding high-yielding drought-tolerant rice: Genetic variations and conventional and molecular approaches. J Exp Bot, 65(21): 6265-6278. [Cited Within:1]
[21]
KumarR, VenuprasadR, AtlinG N. 2007. Genetic analysis of rainfed lowland rice drought tolerance under naturally occurring stress in eastern India: Heritability and QTL effects. Field Crops Res, 103(1): 42-52. [Cited Within:1]
[22]
MackillD J, BonmanJ M. 1992. Inheritance of blast resistance in near isogenic lines of rice. Phytopathology, 82(7): 746-749. [Cited Within:1]
[23]
MishraK K, VikramP, YadawR B, SwamyB P M, DixitS, StaruzM T G, MarkerS, KumarA. 2013. qDTY12. 1: A locus with a consistent effect on grain yield under drought in rice. BMC Genet, 14: 12. [Cited Within:2]
[24]
MurrayH G, ThompsonW F. 1980. Rapid isolation of high molecular weight DNA. Nucl Acids Res, 8(19): 4321-4325. [Cited Within:1]
[25]
MuthuV, AbbaiR, NallathambiJ, RahmanH, RamasamyS, KambaleR, ThulasinathanT, AyyenarB, MuthurajanR. 2020. Pyramiding QTLs controlling tolerance against drought, salinity, and submergence in rice through marker assisted breeding. PLoS One, 15(1): e0227421. [Cited Within:1]
[26]
PatrotiP, VishalakshiB, UmakanthB, SureshJ, SenguttuvelP, MadhavM S. 2019. Marker-assisted pyramiding of major blast resistance genes in Swarna-Sub1, an elite rice variety ( Oryza sativa L. ). Euphytica, 215(11): 179. [Cited Within:2]
[27]
SerrajR, McNallyK L, Slamet-LoedinI, KohliA, HaefeleS M, AtlinG, KumarA. 2011. Drought resistance improvement in rice: An integrated genetic and resource management strategy. Plant Prod Sci, 14(1): 1-14. [Cited Within:1]
[28]
SharmaT R, MadhavM S, SinghB K, ShankerP, JanaT K, DalalV, Pand itA, SinghA, GaikwadK, UpretiH C, SinghN K. 2005. High-resolution mapping, cloning and molecular characterization of the Pi-kh gene of rice, which confers resistance to Magnaporthe grisea. Mol Genet Genomics, 274(6): 569-578. [Cited Within:1]
[29]
SwathiG, DurgaRani C V, JamaloddinM, SheshuMadhav M, VanisreeS, AnuradhaC, RanjitKumar N, ArunaPrem Kumar N, ArunaKumari K, BhogadhiS C, RamprasadE, SravanthiP, KrishinamRaju S, BhuvaneswariV, RajanC P D, JagadeeswarR. 2019. Marker-assisted introgression of the major bacterial blight resistance genes, Xa21 and xa13, and blast resistance gene, Pi54, into the popular rice variety, JGL1798. Mol Breeding, 39: 58. [Cited Within:1]
[30]
VenuprasadR, LafitteH R, AtlinG N. 2007. Response to direct selection for grain yield under drought stress in rice. Crop Sci, 47: 285-293. [Cited Within:1]
[31]
VikramP, SwamyB P M, DixitS, AhmedH U, StaCruz M T, SinghA K, KumarA. 2011. qDTY1. 1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet, 12: 89. [Cited Within:1]
[32]
WassmannR, JagadishS V K, SumflethK, PathakH, HowellG, IsmailA, SerrajR, RedonaE, SinghR K, HeuerS. 2009. Regional vulnerability of climate change impacts on Asian rice production and scope for adaptation. Adv Agron, 102: 91‒133. [Cited Within:1]
1
2019
0.0
0.0
... Anyaoha et al (2019) improved the rainfed rice variety FUNAABOR-2 for grain yield under drought stress by introgressing qDTY12 ...
1
2019
0.0
0.0
... Severe incidence of blast disease was also seen in the upland ecosystem (Asibi et al, 2019), which can be addressed by pyramiding major blast resistance genes ...
2
2007
0.0
0.0
... 1 offers significant yield potential under reproductive-stage drought stress by contributing 51% genetic variance (Bernier et al, 2007) ...
... 1) (Bernier et al, 2007) and blast (two resistance genes Pi54 and Pi1), respectively ...
1
2009
0.0
0.0
... Also, yield is affected due to drought spells in irrigated conditions (Bernier et al, 2009a) ...
1
2009
0.0
0.0
... 1 (Bernier et al, 2009b) ...
1
1973
0.0
0.0
... Milled grains ground to 100 mesh powders were analyzed for gel consistency (Cagampang, 1973) and amylose content (Juliano, 1971) ...
1
2019
0.0
0.0
... Chukwu et al, 2019 ...
2
2012
0.0
0.0
... Rice is also cultivated in around 23 million hectares of rainfed ecosystem where yields are significantly affected due to frequent occurrence of drought (Serraj et al, 2011), and blast (Dean et al, 2012), and global climate change can further increase the drought stress (Wassmann et al, 2009) ...
... Blast disease caused by Magnaporthe grisea (anamorph Pyricularia grisea) is another production constraint commonly observed in the rainfed ecosystem (Dean et al, 2012) ...
1
2015
0.0
0.0
... Devi et al (2015) procedure was used for PCR amplification ...
1
2020
0.0
0.0
... So far, above 100 genes have been reported to aid in blast resistance (Devi et al, 2020) ...
2
2012
0.0
0.0
... 1 A+B region of Dixit et al (2012) showed higher grain yield than the lines possessing either one ...
... 1 B), while RM206 for Pi54, RM224 for Pi1 (Dixit et al, 2012 ...
1
2014
0.0
0.0
... But few reports exist on the development of combined resistant versions to biotic and abiotic stress resistant varieties (Dixit et al, 2014, 2020 ...
2
2020
0.0
0.0
... Marker-assisted breeding strategy has been employed to improve rice varieties for resistance against biotic and biotic stresses independently (Dixit et al, 2020) ...
... But few reports exist on the development of combined resistant versions to biotic and abiotic stress resistant varieties (Dixit et al, 2014, 2020 ...
1
2002
0.0
0.0
... Screening for blastThe blast screening of all the backcross and intercross lines of Varalu was done in a uniform blast nursery at ICAR-IIRR (Umakanth et al, 2017) using Standard Evaluation System with 0‒9 scales (IRRI, 2002) ...
1
2004
0.0
0.0
... Intact milled grains were used to determine grain type by length to width ratio (IRRI, 2004) and alkali spreading value (Juliano et al, 1990) ...
2
2020
0.0
0.0
... Marker-assisted gene pyramiding (MAGP) is a precise, rapid and efficient method to pyramid desired genes/QTLs along with maximum recovery of recurrent parent genome within two to three backcrosses (Jamaloddin et al, 2020) ...
... Like Pi54 and Pi1 pyramided Tellahamsa lines (Jamaloddin et al, 2020), individuals of backcross population having both the genes noted higher resistance score of 0 than the individuals with only Pi54 (0‒2) or Pi1 (2‒3) ...
1
1971
0.0
0.0
... Milled grains ground to 100 mesh powders were analyzed for gel consistency (Cagampang, 1973) and amylose content (Juliano, 1971) ...
1
1990
0.0
0.0
... Intact milled grains were used to determine grain type by length to width ratio (IRRI, 2004) and alkali spreading value (Juliano et al, 1990) ...
1
2018
0.0
0.0
... 5%, respectively, which were higher than the reported introgression studies (Khan et al, 2018 ...
1
2014
0.0
0.0
... Kumar et al, 2014) ...
1
2007
0.0
0.0
... DISCUSSIONSeveral drought-tolerant QTLs detected from diverse sources (Kumar et al, 2007 ...
1
1992
0.0
0.0
... Among them, two significant blast resistance genes Pi54 and Pi1 confer broad-spectrum resistance against predominant races of the pathogen in India (Mackill and Bonman, 1992 ...
2
2013
0.0
0.0
... 1 was evident from the introgressions of Vandana and Sabitri under drought and irrigated conditions (Mishra et al, 2013 ...
... Like Sabitri lines under lowland reproductive stage drought stress (Mishra et al, 2013), the backcross population having properly delimited qDTY12 ...
1
1980
0.0
0.0
... DNA extraction and genotypingGenomic DNA was isolated from 21-day-old seedlings of the parents, F1, backcross and intercross generations (Murray and Thompson, 1980) ...
1
2020
0.0
0.0
... Muthu et al, 2020) ...
2
2019
0.0
0.0
... Thus, Pi54 might provide broad spectrum resistance than Pi1 (Patroti et al, 2019) ...
... Patroti et al, 2019) (Table S1 ...
1
2011
0.0
0.0
... Rice is also cultivated in around 23 million hectares of rainfed ecosystem where yields are significantly affected due to frequent occurrence of drought (Serraj et al, 2011), and blast (Dean et al, 2012), and global climate change can further increase the drought stress (Wassmann et al, 2009) ...
1
2005
0.0
0.0
... Sharma et al, 2005) ...
1
2019
0.0
0.0
... Swathi et al, 2019) ...
1
2007
0.0
0.0
... Screening of derived lines under reproductive stage drought stressBackcross and intercross derived populations were screened for reproductive stage drought stress (Venuprasad et al, 2007) at the research farm, ICAR-Indian Institute of Rice Research (IIRR), Hyderabad, India, in a random block design with 20 cm #cod#x000D7 ...
1
2011
0.0
0.0
... Vikram et al, 2011) are less exploited in breeding programmes ...
1
2009
0.0
0.0
... Rice is also cultivated in around 23 million hectares of rainfed ecosystem where yields are significantly affected due to frequent occurrence of drought (Serraj et al, 2011), and blast (Dean et al, 2012), and global climate change can further increase the drought stress (Wassmann et al, 2009) ...
Improvement of Upland Rice Variety by Pyramiding Drought Tolerance QTL with Two Major Blast Resistance Genes for Sustainable Rice Production








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
 ]
]