


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Functional Analysis of Three Rice Chloroplast Transit Peptides
[He Lei, Chen Guang, Zhang Sen, Qiu Zhennan, Hu Jiang, Zeng Dali, Zhang Guangheng, Dong Guojun, Gao Zhenyu, Ren Deyong, Shen Lan, Guo Longbiao, Qian Qian , Zhu Li]
, Zhu Li]
, Zhu Li]
|
|

 , Zhu Li
, Zhu Li
Chloroplast transit peptides (CTPs) can be used to transport non-chloroplastic proteins into the chloroplasts. Here, we studied the CTPs of three rice ( Oryza sativa L.) chloroplast-localized proteins and found that their CTPs could be used to transport non-chloroplast-localized proteins into the chloroplasts. Fusion proteins lacking the CTP remained located in the cytoplasm. Furthermore, we constructed green fluorescent protein fusion vectors with the three CTPs and three non-chloroplast-localized proteins, Ghd10, MULTI-FLORET SPIKELET1 (MFS1), and SHORTENED UPPERMOST INTERNODE 1 (SUI1). After transforming these constructs into rice protoplasts, the fusion proteins all localized in the chloroplasts. Collectively, our results showed that these CTPs can transport non-chloroplast-localized proteins into the chloroplasts, and more importantly, these CTPs can be applied to engineer chloroplast metabolism.
In addition to their essential roles in photosynthesis and CO2 fixation, chloroplasts participate in the synthesis of chlorophyll, carotenoids, fatty acids, some amino acids, starch and proteins (Hanke and Mulo, 2013). The chloroplast genome encodes only around 100 proteins (Martin et al, 2002; Timmis et al, 2004; Tu et al, 2017), and therefore, a large number of proteins that function in the chloroplasts are synthesized in the cytoplasm (Leister, 2003; Sun et al, 2009), then transported into the chloroplast and processed into mature proteins. This transport is usually mediated by the chloroplast transit peptide (CTP) in the precursor sequence of the protein, a sequence that is post-translationally cleaved by stromal processing peptidase in the chloroplast stroma (Keegstra and Bauerle, 1988; von Heijne et al, 1989). Many studies have shown that CTPs mediate the transportation of precursor proteins into chloroplasts via interaction with certain translocators, such as Hsp70 (heat shock proteins 70) (Ivey and Bruce, 2000), Hsp90 (heat shock proteins 90) (Qbadou et al, 2006), FKBP (FK506 binding protein) (Fellerer et al, 2011), TIC (translocon at the inner membrane of chloroplasts) and TOC (translocon at the outer membrane of chloroplasts) complexes (Sun et al, 2011), which are located at the inner and outer envelopes of chloroplasts.
CTPs mediate the transport of chloroplast proteins from the cytoplasm into the chloroplast and can be used to transport non-chloroplast proteins into the chloroplast. In 1985, van denBroeck et al (1985) successfully transported bacterial neomycin phosphor- transferase II (NPT-II) into tobacco chloroplasts by ligating a 58-amino acid transit peptide of the pea (Pisum sativum) Ribulose-1, 5-bisphosphate carboxylase/ oxygenase (Rubisco) small subunit to the N-terminal of NPT-II. Exogenous disease and insect ré sistance genes are transformed to chloroplast by using chloroplast transit peptide, which enhances the ability of transgenic plants to resist diseases and insects. Li (2016) transported yeast acyl-δ 9 dehydrogenase into tobacco (Nicotiana tabacum) chloroplasts by using a CTP from Arabidopsis thaliana Ribulose-1, 5-bisphosphate carboxylase/oxygenase (Rubisco) small subunit and found that the effect of this enzyme on fatty acid composition in the chloroplasts is stronger than that in the cytoplasm.
The CTP of 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) in Sorghum bicolor is fused to the EPSPS of Agrobacterium tumefaciens and transformed into maize (Zea mays), which showed that the CTP plays an important role in glyphosate resistance conferred by EPSPS. When the chloroplast transit peptide is absent, although EPSPS is still highly expressed, the plant possess no glyphosate resistance (Zhao et al, 2013). Li et al (2013) fused the Rubisco CTP from rice with β -glucuronidase (GUS) and transformed it into rice, and found that the rice plants with Rubisco CTP increases GUS protein expression levels for about 4.2-fold, compared to the plants without CTP.
Accumulating evidence showed that bioengineering technologies are important to solve the world food problem by improving the photosynthesis, stress resistance and yield traits of crop (Long et al, 2006; Zhang, 2007; Li et al, 2013; Zhao et al, 2013; Shen et al, 2017). For the biotechnology process, exogenous proteins often need to be effectively introduced into chloroplasts, so it is very important for CTP to work well (Shen et al, 2017). Although many studies have been done to research the function of chloroplast transit peptides in plants, most of the related studies in CTP use the Rubisco small subunit transit peptide for chloroplastic transportation (von Heijne et al, 1989; Wong et al, 1992; Leister, 2003; Kim et al, 2010; Fellerer et al, 2011; Sun et al, 2011; Li et al, 2013; Zhao et al, 2013). Therefore, more diverse transit peptides are required for genetic engineering, especially for metabolic engineering (Wong et al, 1992; Kim et al, 2010; Li et al, 2013).
Recently, we isolated and identified a rice albino mutant, hsa1, which is sensitive to heat stress. The results of map-based cloning showed that HSA1 encodes a putative FLN2 (fructokinase-like protein 2) protein. Further protein homology analysis suggested that the rice genome contains genes encoding two FLN isoforms, FLN1 (LOC_Os01g63220) and FLN2/HSA1 (LOC_Os03g40550) (Qiu et al, 2018). Using yeast two-hybrid assays, we showed that OsFLN1 and HSA1/OsFLN2 interacted with OsTRXz (thioredoxin protein z), and fln1 and trxz mutant alleles, created via Clustered Regularly Interspaced Short Palindromic Repeats (CRISPR)/CRISPR-associated protein 9 (Cas9), were associated with a severe albino phenotype and seedling lethality. Our results showed that FLN1 and HSA1 interact with TRXz (LOC_Os08g29110) to regulate chloroplast biogenesis and plant growth (He et al, 2018). In this study, we used bioinformatics to predict the CTP sequence of TRXz, FLN1 and HSA1. To verify the accuracy of our predicted CTPs, we observed the subcellular localization of the three proteins without their CTPs. Then, we investigated the function of CTP from FLN1, HSA1 and TRXz, and demonstrated that this CTP could localize a non-chloroplast protein into the chloroplast. The discovery of the sequence and function of these CTPs provided insight into plant stress responses and a potentially useful tool for chloroplast metabolic engineering.
For protoplast isolation, Nipponbare (NPB, Oryza sativaL. japonica) seeds were sown on Murashige and Skoog (MS) medium (Murashige and Skoog, 1962) and cultured under 16 h light / 8 h dark photoperiods at 28 º C under illumination with 200 μ mol/(m2∙ s). The p35S::GFP vector was obtained from our laboratory. The reagents for protoplast isolation and transformation were purchased from Sigma- Aldrich (Shanghai, China).
We selected three chloroplast proteins, FLN1, TRXz, and HSA1/FLN2 (He et al, 2018). To further clarify the function of the CTPs of these three proteins, we obtained their protein sequences from the Rice Genome Annotation Project Resource website (http://rice.plantbiology.msu.edu/) and used the TargetP website (http://www.cbs.dtu.dk/services/TargetP/) to predict the sequence of their CTPs.
The p35S::GFP vector was digested with SalI (Takara Biotech, Dalian, China) at 37 º C for 2 h in a 50 μ L reaction, consisting of 5 μ L of 10× H buffer, 10 μ L p35S::GFP vector, 2 μ L SalI, and 33 μ L sterile water. The digested products were analyzed on 1% agarose gels. We used the Wizard SV Gel and PCR Clean-Up System (Promega Biotech, USA) to extract and purify the target fragment.
To construct the p35S::CTPTRXz-GFP vector, we generated a DNA fragment encoding the N-terminal 56 amino acids of TRXz by PCR using p35S:: CTPTRXz-GFP primer (Table 1) from the NPB cDNA. The PCR procedure was as follows: Denaturation at 94 º C for 4 min, followed by 32 cycles of 98 º C for 10 s, annealing at 55 º C for 30 s, extension at 68 º C for 30 s, and a final extension step at 68 º C for 10 min. The PCR products were analyzed on 2% agarose gels. We used the Wizard SV Gel and PCR Clean-Up System (Promega Biotech, USA) to extract and purify the target DNA fragment. The purified DNA fragment encoding the N-terminal 56 amino acids of TRXz was fused with the purified p35S::GFP fragment using a ClonExpress II One Step Cloning Kit (Vazyme Biotech, Nanjing, China) by homologous recombination. The ligation reaction was conducted for 30 min at 37 º C in a 20 μ L reaction, consisting of 2 μ L PCR product, 2 μ L purified p35S::GFP fragment, 4 μ L of 5× CEII buffer, 2 μ L Exnase II and 10 μ L sterile water. After transformation intoEscherichia coli (DH5α ), the positive clones were identified by PCR using the s65t-1F (GAGGACAGGCTTCTTGAG) and s65tR (GGTGGTGCAGATGAACTT) primers and sent to Hangzhou Tsingke Biotech Company (Hangzhou, China) for sequencing verification using the s65t-1F and s65tR primers. The other CTP and GFP fusion protein vectors (p35S::CTPHSA1-GFP and p35S:: 36CTPFLN1-GFP) were constructed and verified as described above using primers listed in Table 1.
| Table 1 Nucleotide sequences used in construction of fusion vector with chloroplast transit peptides and GFP (green fluorescent protein). |
To construct CTP-deleted proteins and GFP fusion vectors, we generated DNA fragments encoding deletion of CTP of TRXz and FLN1 by PCR using primers listed in Table 2 from NPB cDNA respectively. The PCR procedure, ligation reaction and positive clones verification was as above.
| Table 2 Nucleotide sequences used in construction of GFP (green fluorescent protein) fusion vector for deletion of chloroplast transit peptide. |
To construct the p35S::CTPTRXz-Ghd10-GFP vector, we generated a DNA fragment encoding the N-terminal 56 amino acids of TRXz by PCR using p35S::CTPTRXz-Ghd10-GFP-1 primer (Table 3) from the NPB cDNA and generated the DNA fragment encoding Ghd10 by PCR using p35S::CTPTRXz-Ghd10- GFP-2 primer (Table 3) from the NPB cDNA. The PCR procedure was as above. The purified DNA fragments encoding the N-terminal 56 amino acids of TRXz and Ghd10 were fused with the purified p35S::GFP fragment using a ClonExpress MultiS One Step Cloning Kit (Vazyme Biotech, Nanjing, China) by homologous recombination. The ligation reaction was conducted for 30 min at 37 º C in a 20 μ L reaction, consisting of 2 μ L each PCR product, 4 μ L purified p35S::GFP fragment, 4 μ L of 5× CE MultiS buffer, 2 μ L Exnase MultiS, and 6 μ L sterile water. After transformation into Escherichia coli(DH5α ), the positive clones were identified by PCR and sequence verification using the s65t-1F and s65tR primers. The other vectors were constructed and verified as described above using primers listed in Table 3.
| Table 3 Nucleotide sequences used in construction of GFP fusion vector with non-chloroplast-localized gene and chloroplast transit peptide. |
Protoplasts were isolated from 2- to 3-week-old rice seedlings according to Wu et al (2016). Briefly, about 100 stems and sheaths of rice were cut into 0.5 mm pieces and digested in 15 mL enzyme solution (1.5% cellulase R-10, 0.75% macerozyme R-10, 0.6 mol/L mannitol, 10 mmol/L 2-(N-Morpholino)ethanesulfonic acid hydrate with pH 5.7, 10 mmol/L CaCl2, and 0.1% bovine serum albumin) for 6 h at 28 º C. After filtering through a piece of miracloth, protoplasts were pelleted by a 5-min centrifugation at 80 × g in a horizontal centrifuge and resuspended in 10 mL W5 solution (154 mmol/L NaCl, 125 mmol/L CaCl2, 5 mmol/L KCl, and 2 mmol/L MES with pH 5.7). After resting on ice for at least 30 min, protoplasts were spun down by another 1 min centrifugation at 100 × g and resuspended into 2 mL MMG solution (0.4 mol/L mannitol, 15 mmol/L MgCl2, and 4 mmol/L MES with pH 5.7). The protoplasts were counted using a hemocytometer under a microscope. The protoplasts were diluted into the optimal working concentration of 2 × 105 cells/mL.
DNA transfection was carried out in a 2-mL round bottom microcentrifuge tube, in which 100 μ L protoplasts (2 × 104 cells) were mixed well with 20 μ g (about 20 μ L) constructs. Then, 120 μ L PEG solution (40% PEG4000, 0.2 mol/L mannitol, and 0.1 mol/L CaCl2) was added to each tube, and transfection was initiated sequentially by a gentle tapping of the tube bottom 15 times. After a 15-min incubation at 28 º C, transfection was terminated in the same order by adding 1000 μ L W5 solution and inverting the tube gently three times. The solution was centrifuged at 80 × g for 2 min, and the supernatant was discarded. Then, 2 mL W5 solution was added to gently resuspend the protoplast pellet, and protoplasts were cultured for 16-20 h in darkness to allow expression of the GFP fusion proteins. Before confocal microscopy, the transformed protoplasts were centrifuged at 80 × g for 2 min to remove most of the supernatant. GFP fluorescence was detected by a confocal laser- scanning microscopy (Olympus IX71, Tokyo, Japan).
FLN1, HSA1 and TRXz localize in the chloroplast. To clarify the function of the CTPs of these three proteins, we predicted their CTP sequences with TargetP (http:// www.cbs.dtu.dk/services/TargetP/). Protein sequence analysis using TargetP suggested that the 36 amino acids of the FLN1 N-terminal region formed a CTP, whereas the 54 amino acids of the HSA1 N-terminal region, and the 56 amino acids of the TRXz N-terminal region formed a CTP, respectively (Fig. 1).
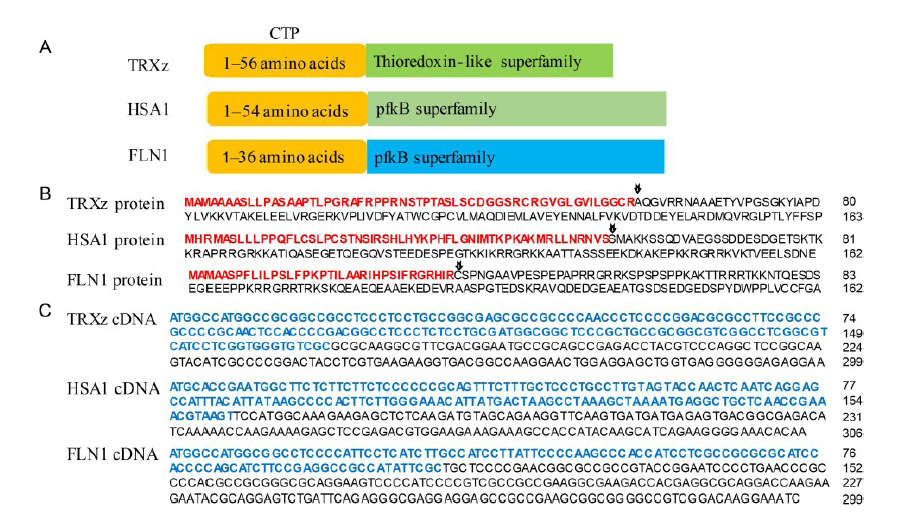 | Fig. 1. Structure and sequence of FLN1, HSA1 and TRXz. A, Abbreviated diagram of TRXz, HSA1 and FLN1 protein sequences. B, Amino acids sequences of the three proteins. Sequences of the chloroplast transit peptide (CTP) are shown in red bold type. The arrows indicate the putative cleavage site during chloroplast import based on the TargetP website. C, cDNA sequences encoding the three genes. Sequences of the three CTPs are shown in blue bold type. |
In our previous study, we found that TRXz was located in the chloroplast (He et al, 2018). TargetP predicted that the N-terminal 56 amino acids of TRXz is the CTP (CTPTRXz). To verify this, we constructed the p35S::CTPTRXz-GFP vector containing a DNA fragment encoding the N-terminal 56 amino acids of TRXz (Fig. 2-A) and the p35S::Δ CTPTRXz-GFP vector containing a DNA fragment encoding 133 amino acids of TRXz that lacks the N-terminal 56 amino acids (Δ CTPTRXz) (Fig. 2-B). We transformed these two vectors into rice protoplasts and found that the Δ CTPTRXz-GFP fusion protein was located in the cytoplasm (Fig. 2-G), and the CTPTRXz-GFP fusion protein was located in the chloroplast (Fig. 2-H).
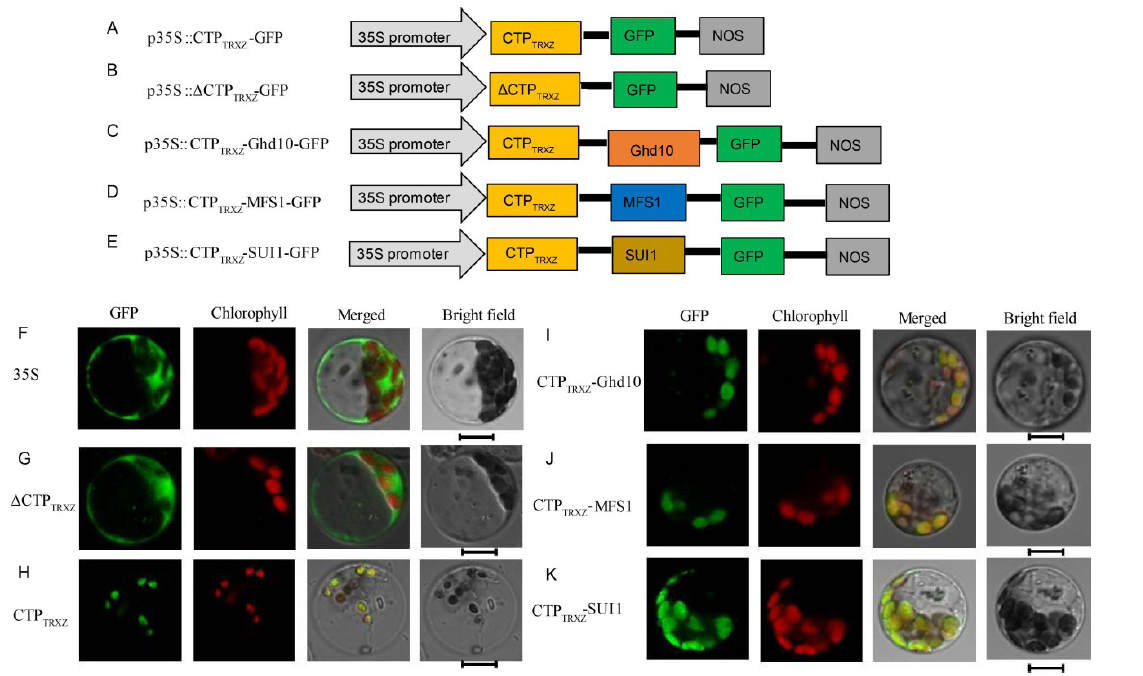 | Fig. 2. Subcellular localization of TRXz and other proteins. A-E, Diagrams of vector construction; F, The free green fluorescent protein (GFP) protein as a control. G, The Δ CTPTRXz protein localized to the cytoplasm. H, The only N-terminal 56 amino acids of the TRXz and GFP fusion protein localized to chloroplast. I-K, The CTPTrxz-Ghd10, CTPTrxz-MFS1 and CTPTrxz-SUI1 fusion proteins localized to the chloroplasts. All of the GFP fusion proteins carried a C-terminal GFP tag. In F-K, GFP fluorescence, chlorophyll autofluorescence, merged GFP and chlorophyll fluorescence, and bright-field images are shown. Bar = 5 μ m. |
To further investigate whether the CTP can transport non-chloroplast-localized proteins into the chloroplast, we selected three proteins: Ghd10 (LOC_Os10g28330), MFS1 (LOC_Os05g41760) and SUI1 (LOC_Os01g02890). SUI1 is located in the nuclear membrane and cell membrane (Zhu et al, 2011; Zhao et al, 2015), whereas Ghd10 and MFS1 are located in the nucleus (Hu et al, 2013; Ren et al, 2013). We constructed the p35S::CTPTRXz-Ghd10-GFP vector containing a DNA fragment encoding the N-terminal 56 amino acids of TRXz and a full-length DNA segment of Ghd10 (Fig. 2-C), p35S::CTPTRXz-MFS1- GFP containing a DNA fragment encoding the N-terminal 56 amino acids of TRXz and a full-length DNA segment of MFS1 (Fig. 2-D), and p35S::CTPTRXz- SUI1-GFP containing a DNA fragment encoding the N-terminal 56 amino acids of TRXz and a full-length DNA segment of SUI1 (Fig. 2-E). We introduced these vectors into rice protoplasts. Green fluorescence showed that all the three fusion proteins localized in the chloroplasts (Fig. 2-I to -K). These results suggested that the N-terminal 56 amino acids of TRXz comprise its CTP, which functions to guide the protein into the chloroplasts.
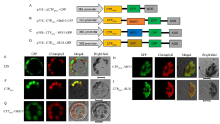
HSA1 localized in the chloroplasts and the Δ CTPHSA1-GFP fusion protein localized in the cytoplasm (Qiu et al, 2018). To determine whether the N-terminal 54 amino acids comprise its CTP, we constructed the p35S::CTPHSA1-GFP vector containing a DNA fragment encoding the N-terminal 54 amino acids of HSA1 (CTPHSA1, Fig. 3-A). We transformed the vector into rice protoplasts and found that the CTPHSA1-GFP fusion protein localized in the chloroplasts (Fig. 3-F).
We constructed the p35S::CTPHSA1-Ghd10-GFP vector containing a DNA fragment encoding the N-terminal 54 amino acids of HSA1 and a full-length DNA segment of Ghd10 (Fig. 3-B), p35S::CTPHSA1- MFS1-GFP containing a DNA fragment encoding the N-terminal 54 amino acids of HSA1 and a full-length DNA segment of MFS1 (Fig. 3-C), and p35S::CTPHSA1- SUI1-GFP containing a DNA fragment encoding the N-terminal 54 amino acids of HSA1 and a full-length DNA segment of SUI1 (Fig. 3-D). We introduced these vectors into rice protoplasts. Green fluorescence showed that all the three fusion proteins localized in the chloroplasts (Fig. 3-G to -I). Taken together, these results indicated that the N-terminal 54 amino acids of HSA1 comprise its CTP, which can transport other non-chloroplast proteins into the chloroplasts.
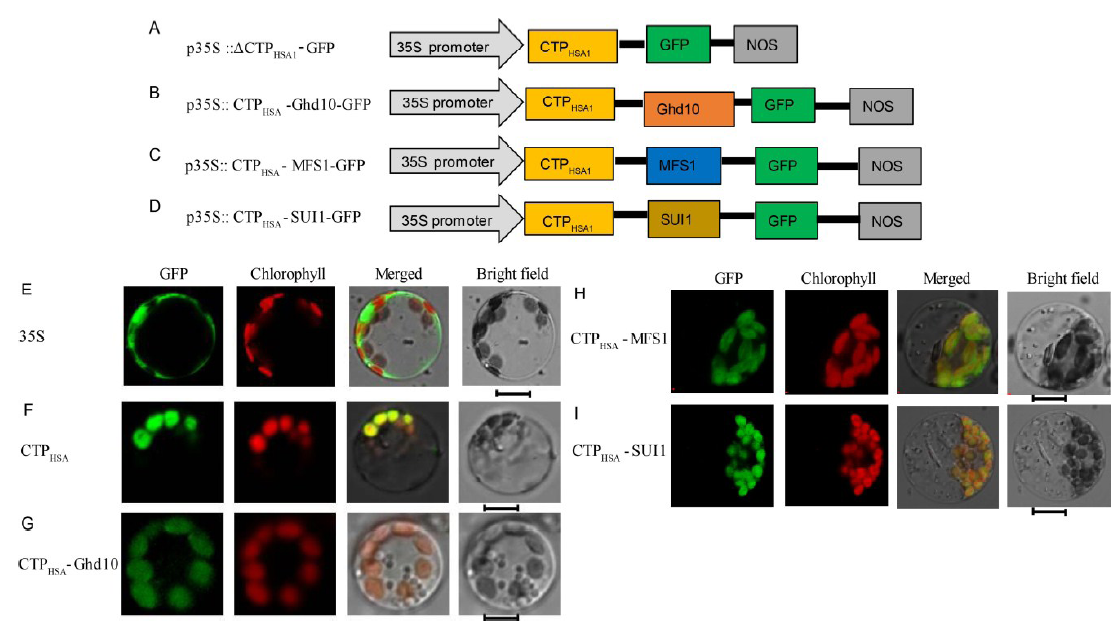 | Fig. 3. Subcellular localization of HSA1 and other proteins. A-D, Diagrams of vector construction. E, The free green fluorescent protein (GFP) protein as a control. F, The only N-terminal 54 amino acids of the HSA1 and GFP fusion protein localized to chloroplast. G-I, The CTPHSA1-Ghd10, CTPHSA1-MFS1 and CTPHSA1-SUI1 fusion proteins localized to the chloroplasts. All of the GFP fusion proteins carried a C-terminal GFP tag. In E-I, GFP fluorescence, chlorophyll auto fluorescence, merged GFP and chlorophyll fluorescence, and bright-field images are shown. Bar = 5 μ m. |
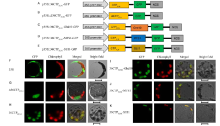
The HSA1/FLN2 homologous protein FLN1 localizes in the chloroplasts (Qiu et al, 2018). TargetP predicted that the N-terminal 36 amino acids comprise its CTP (36CTPFLN1). To verify this, we constructed the p35S:: 36CTPFLN1-GFP vector containing a DNA fragment encoding the N-terminal 36 amino acids of FLN1 (Fig. 4-A) and the p35S::Δ 36CTPFLN1-GFP vector containing a DNA fragment encoding 495 amino acids of FLN1 that lacks the N-terminal 36 amino acids (Δ 36CTPFLN1, Fig. 4-B). We transformed these two vectors into rice protoplasts and found that the Δ 36CTPFLN1-GFP fusion protein localized in the cytoplasm (Fig. 4-G) and 36CTPFLN1-GFP fusion protein localized in the chloroplasts (Fig. 4-H).
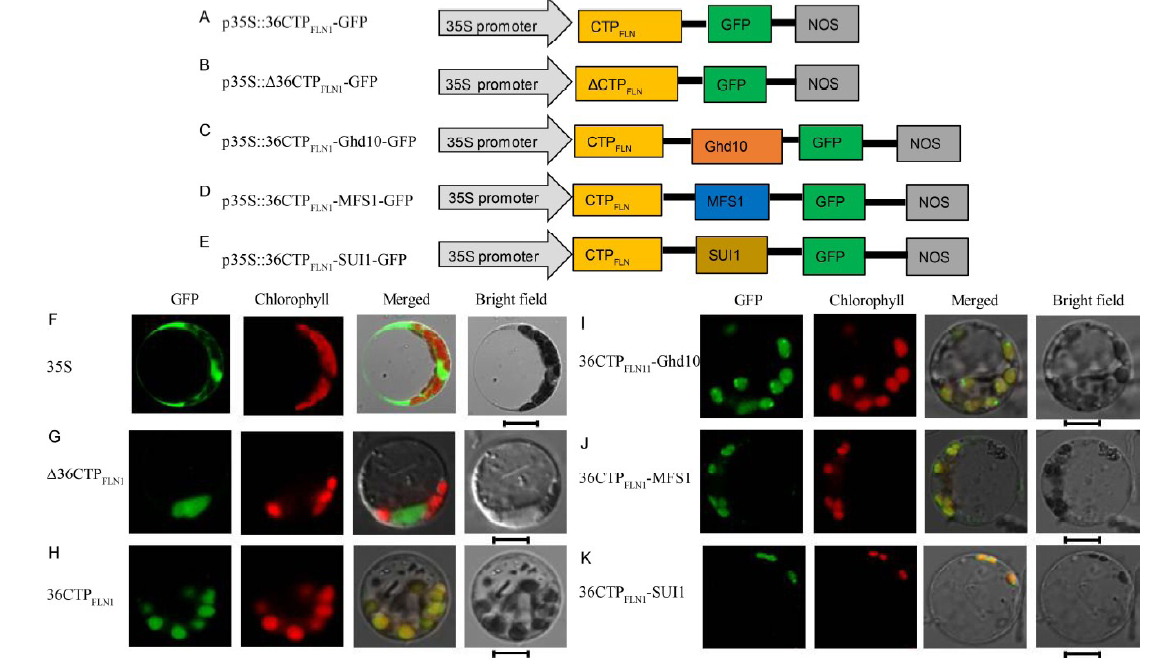 | Fig. 4. Subcellular localization of FLN1 and other proteins. A-E, Diagrams of vector construction. F, The free green fluorescent protein (GFP) protein as a control. G, The Δ 36CTPFLN1 protein localized to the cytoplasm. H, The only N-terminal 36 amino acids of the FLN1 and GFP fusion protein localized to chloroplast. I-K, The 36CTPFLN1-Ghd10, 36CTPFLN1-MFS1, and 36CTPFLN1-SUI1 fusion proteins localized to the chloroplasts. All of the GFP fusion proteins carried a C-terminal GFP tag. In F-K, GFP fluorescence, chlorophyll auto fluorescence, merged GFP and chlorophyll autofluorescence, and bright-field images are shown. Bar = 5 μ m. |
We constructed the p35S::36CTPFLN1-Ghd10-GFP vector containing a DNA fragment encoding the N-terminal 36 amino acids of FLN1 and a full-length DNA segment of Ghd10 (Fig. 4-C); p35S::36CTPFLN1- MFS1-GFP containing a DNA fragment encoding the N-terminal 36 amino acids of FLN1 and a full-length DNA segment of MFS1 (Fig. 4-D); and p35S:: 36CTPFLN1-SUI1-GFP containing a DNA fragment encoding the N terminal 36 amino acids of FLN1 and a full-length DNA segment of SUI1 (Fig. 4-E). We introduced these vectors into rice protoplasts. Green fluorescence showed that all the three fusion proteins localized in the chloroplasts (Fig. 4-I to -K). These results showed that the N terminal 36 amino acids of FLN1 are necessary for the transport of FLN1 into the chloroplasts and can also transport non-chloroplast proteins into the chloroplasts.
Our results showed that the Δ CTPTRXz-GFP and Δ 36CTPFLN1-GFP fusion proteins were not localized in the chloroplasts (Figs. 2-G and 4-G), whereas the GFP signals of CTPTRXz-GFP, CTPHSA1-GFP and 36CTPFLN1-GFP fusion proteins were observed to be concomitant with the chlorophyll autofluorescence (Figs. 2-H, 3-F and 4-H). These results suggested that the N-terminal 56 amino acids of TRXz, the N-terminal 54 amino acids of HSA1, and the N-terminal 36 amino acids of FLN1 are their respective CTPs. To further verify the function of these CTPs, we fused the CTPs with non-chloroplast proteins, SUI1, Ghd10 and MFS1, tagged with GFP. After transformation of these fusion proteins into rice protoplasts, the green fluorescence signal was found in the chloroplasts (Figs. 2-I to -K, 3-G to -I and 4-I to -K). These results indicated that these CTPs not only guide the chloroplast proteins to the chloroplasts, but also guide foreign proteins to the chloroplasts.
The prediction of CTP sequences is based on the existing sequence information, which may contain errors. In this study, we found that after removing the 36 amino acids from the N-terminal of FLN1, the green fluorescence signal was not observed throughout the cytoplasm and cell membrane, but only in a small area of the cytoplasm (Fig. 4-G). We constructed GFP fusion vectors containing a DNA fragment encoding 481 or 442 amino acids of FLN1 that lacks the N-terminal 50 or 89 amino acids of FLN1 (Δ 50CTPFLN1 and Δ 89CTPFLN1) p35S:: Δ 50CTPFLN1-GFP and p35S::Δ 89CTPFLN1-GFP (Fig. 5-A and -B). After transformation into rice protoplasts, we found that p35S::Δ 50CTPFLN1-GFP vector still existed in a small region of the cytoplasm, while the fluorescence signal of the transformed p35S:: Δ 89CTPFLN1-GFP vector filled the cytoplasm and cell membrane (Fig. 5-F and -G). Furthermore, we constructed two fusion vectors p35S::50CTPFLN1- SUI1-GFP and p35S::89CTPFLN1-SUI1-GFP, containing a DNA fragment encoding the N terminal 50 or 89 amino acids of FLN1 (Fig. 5-C and -D), and a full-length segment of SUI1 tagged with GFP and transformed them into rice protoplasts. The fusion proteins, containing the N-terminal 50 or 89 amino acids and SUI1-GFP, localized in the chloroplasts (Fig. 5-H and -I). These results suggest that N-terminal 89 amino acids of FLN1 comprise its CTP.
Transferring foreign proteins into chloroplasts can change photosynthetic characteristics. Currently, the most commonly used approach is to guide the foreign protein into the chloroplasts by use of a CTP, which is typically fused to the N-terminus of the target protein. The Rubisco small subunit CTP is the most commonly used CTP for targeting foreign proteins into chloroplasts (Wong et al, 1992; Zhong et al, 2003; Kim et al, 2010). In this study, we reported three new CTPs in rice. Transient expression analysis in rice protoplasts demonstrated that these three CTPs can transport foreign proteins into the chloroplasts. These CTPs may be useful as targets for chloroplast metabolic engineering, particularly for crop plants.
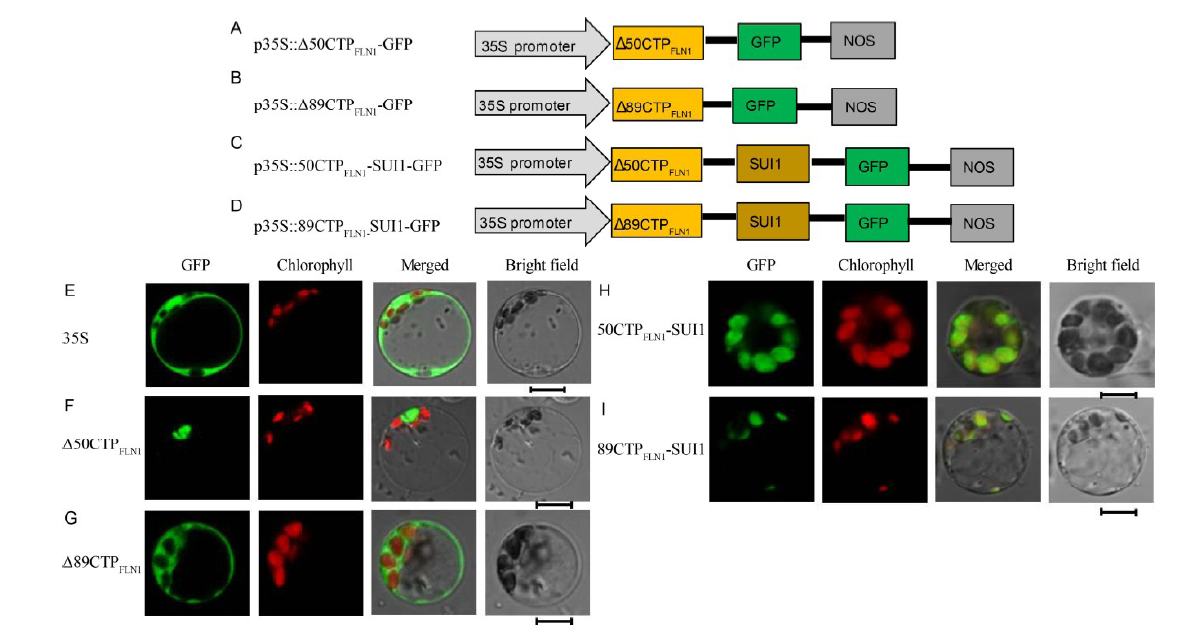 | Fig. 5. Subcellular localization of other FLN1-related proteins. A-D, Diagrams of vector construction. E, The free green fluorescent protein (GFP) protein as a control. F, The Δ 50CTPFLN1 protein localized to the cytoplasm. G, The Δ 89CTPFLN1 protein localized to the cytoplasm. H and I, The 50CTPFLN1-SUI1 and 89CTPFLN1-SUI1 fusion proteins localized to the chloroplasts. All of the GFP fusion proteins carried a C-terminal GFP tag. In E-I, GFP fluorescence, chlorophyll autofluorescence, merged GFP and chlorophyll fluorescence, and bright-field images are shown. Bar = 5 μ m. |
The work was supported by the National Natural Science Foundation of China (Grant Nos. 31601284 and 31661143006), the Transgenic Plant Research and Commercialization Project of the Ministry of Agriculture of China (Grant No. 2016ZX08001003-002), Zhejiang Province Outstanding Youth Fund (Grant No. LR16C130001), and the Collaborative Innovation Project of the Chinese Academy of Agricultural Sciences (Grant No. Y2016XT05).
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|